H-Evolution-005Eukaryotic
Eukayotes
First Eukayotes: 1.7 billion years ago Eukayotic cells came into being. These big cells by comparison seem to have captured the small cells with special capability and enslaved them, and eventually encompass them as an integral part of the bacterial cell colony encased in a membrane. This cleaver membrane is a double layer of molecule soldiers, one layer opens and closes before the other layer opens and closes, thus letting waste out and bringing food in.


Prior to the electron microscope, humans were oblivious to the existence of small things.
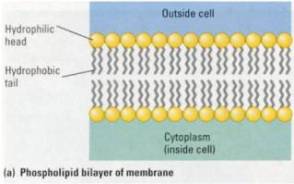
Perimeter “storm door” cell walls
Protists
“No more pleasant
sight has met my eye than this of so many thousands of creatures in one small
drop of water” wrote Anton van Leeuwenhoek after his discovery of the microbial
world more than three centuries ago. It a world every biology student should
have the opportunity to rediscover by peering through a microscope into a
droplet of pond water filled with diverse creatures we call protists. Protists
are eukaryotic, and thus even the simplest are much more complex than the
prokaryotes. The first eukaryotes to evolve from prokaryotic ancestors were
protists. The very word implies great antiquity (from the Greek protos, first)
.The primal eukaryotes were not only the predecessors of the great variety of
modern protists, but were also ancestral to all other eukaryotes-plants, fungi,
and animals. Two of the most significant chapters in the history of life-the
origin of the eukaryotic cell and the subsequent emergence of multicellular
eukaryotes-unfolded during the evolution of protists.
The
Origin of Eukaryotic Cells .
The many
differences between prokaryotic and eukaryotic cells far outnumber the
differences between plant and animal cells. The fossil record indicates that
eukaryotes evolved from prokaryotes more than 1.7 billion years ago. One of
biology's most engaging questions is how this happened-in particular, how the
membrane-enclosed organelles of eukaryotic cells arose. A widely accepted
theory is that eukaryotic cells evolved through a combination of two processes.
In one process, the eukaryotic cell's endomembrane system-all the
membrane-enclosed organelles except mitochondria and chloroplasts evolved from
inward folds of the plasma membrane of a prokaryotic cell (1>2) A second, very different process, called
endosymbiosis, generated mitochondria and chloroplasts.(3>4 & 5>6)

Figure 15.18 How did
eukaryote cells evolve?
Symbiosis is a
close association between organisms of two or more species. The word symbiosis
is from the Greek for "living together” and endosymbiosis refers to one
species living within another, called the host. chloroplasts and mitochondria
evolved from small symbiotic prokaryotes that established residence within
other, larger host prokaryotes (Figure 15.18b ). The ancestors of mitochondria
may have been aerobic bacteria that were able to use oxygen to release large
amounts of energy from organic molecules by cellular respiration. At some
point, such a prokaryote might have been an internal parasite of a larger
heterotroph, or an ancestral host cell may have ingested some of these aerobic
cells for food. If some of the smaller cells were indigestible, they might have
remained alive and continued to perform respiration in the host cell. In a
similar way, photosynthetic bacteria ancestral to chloroplasts may have come to
live inside a larger host cell. Because almost all eukaryotes have mitochondria
but only some have chloroplasts, it is likely that mitochondria evolved first.
By whatever means the relationships
began, it is not hard to imagine the symbiosis eventually becoming mutually
beneficial. In a world that was becoming increasingly aerobic, a cell that was
itself an anaerobe would have benefited from aerobic endosymbionts that turned
the oxygen to advantage. And a heterotrophic host could derive nourishment from
photosynthetic endosymbionts In the process of becoming more interdependent,
the host and endosymbionts would have become a single organism, its parts
inseparable.
Developed most
extensively by Lynn Margulis, the endosymbiosis theory is supported by
extensive evidence. Present-day mitochondria and chloroplasts are similar to
prokaryotic cells in a number of ways. For example, both types of organelles
contain small amounts of DNA, RNA, and ribosomes that resemble prokaryotic
versions more than eukaryotic ones. These components enable cWoroplasts and
mitochondria to exhibit some autonomy in their activities. The organelles
transcribe and translate their DNA into polypeptides, contributing to some of
their own enzymes. They also replicate their own DNA and reproduce within the
cell by a process resembling the binary fission of prokaryotes.
The origin of the
eukaryotic cell made more complex organisms possible, and a vast variety of
protists evolved.
The
Diversity of Protists
All protists are
eukaryotes, but they are so diverse that few other general characteristics can
be cited. In fact, protists vary in structure and function more than any other
group of organisms. Most protists are unicellular, but there are some colonial
and multicellular species. Because most protists are unicellular, they are
justifiably considered the simplest eukaryotic organisms. But at the cellular
level, many protists are exceedingly complex-the most elaborate of all cells.
We should expect this of organisms that must carry out within the boundaries of
a single cell, all the basic functions performed by the collective of
specialized cells that make up the bodies of plants and animals. Each
unicellular protist is not at all analogous to a single cell from a human, but
is itself an organism as complete as any whole animal or plant.
For our survey of
these diverse organisms, we'll look at four major categories of protists,
grouped-more by lifestyle than by their evolutionary relationships: protozoans,
slime molds, unicellular algae, and seaweeds.
Protozoans
Protists that live primarily by ingesting food, a mode of
nutrition that is animal-Iike, are called protozoans ("first
animal"). Protozoans thrive in all types of aquatic environments,
including wet soil and the watery environment inside animals. Most species eat
bacteria or other protozoans, but some can absorb nutrients dissolved in the
water. Protozoans that live as parasites in animals, though in the minority,
cause some of the world's most harmful human diseases. We'll examine five
groups of protozoans: flagellates, amoebas, forams, apicomplexans, and
ciliates.
Flagellates
are protozoans that move by means of one or more flagella. Most species are
free-Iiving (nonparasitic). However, there are also some nasty parasites that
make humans sick. An example is Giardia, a flagellate that infects the human
intestine and can cause abdominal cramps and severe diarrhea. People become
infected mainly by drinking water contaminated with feces from infected
animals. Giardia can ruin a camping trip. Another group of dangerous
flagellates are the trypanosomes, including a species that causes sleeping
sickness, a serious illness prevalent in tropical Africa and transmitted by the
tsetse fly (Figure 15.19a)
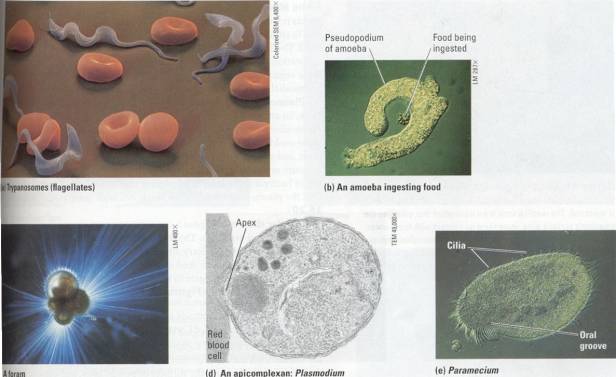
Figure 15.19 Examples of protozoans.(a) Trypanosomes are
flagellates that live as parasites in the bloodstream of vertebrate animals.
The squiggles among these human red blood cells are trypanosomes that cause
sleeping sickness, a debilitating disease common in parts of Africa. Trypanosomes escape being killed by their
host's defenses by being quick-change artists. They alter the molecular
structure of their coats frequently, thus preventing immunity from developing
in the host. (b) This amoeba is
ingesting a smaller protozoan as food. The amoeba's pseuilopodia arch around
the prey and engulf it into a food vacuole (also see Chapter 5). (c) Forams are
almost all marine. The foram cell secretes a porous, multichambered shell made
of organic material hardened with calcium carbonate, the same mineral that
makes up limestone. Thin strands of cytoplasm (pseudopodia) extend through the
pores, functioning in swimming, shell formation, and feeding. The shells of
fossilized forams are major components of the limestone rocks that are now land
formations. (d) Plasmodium, the apicomplexan that causes malaria, uses its
apical complex to enter red blood cells of its human host. The parasite feeds
on the host cell from within, eventually destroying it. (e) The ciliate
Paramecium uses its cilia to move through pond water. Cilia also line an
indentation called the oral groove, and their beating keeps a current of water
containing bacteria and small protists moving toward the cell "mouth"
at the base of the groove.
Amoebas are characterized by
great flexibility and the absence of permanent locomotor organelles. Most
species move and feed by means of pseudopodia ( singular, pseudopodium )',
temporary extensions of the cell (Figure 15.19b) .Amoebas can assume virtually
any shape as they creep over rocks,
sticks, or mud at the bottoII1 of a pond or ocean. Other protoroans with
pseudopodia include the forams (Figure 15.19c).
Apicomplexans
are all parasitic, and some cause serious human diseases. Theyare named for an
apparatus at their apex that is specialized for penetrating host cells and
tissues. This protozoan group includes Plasmodium, dIe parasite that causes
malaria (Figure 15.19d). Spread by mosquitoes, malaria is one of the most
debilitating and widespread human diseases. Each year in the tropics, more than
200 million people become infected, and at least a million die in Africa alone.
As part of the effort to combat malaria, scientists determined the complete
sequence of the Plasmodium genome in 2002.
Ciliates are
protozoans that use locomotor structures called cilia to move and feed.
Nearlyall ciliates are free-living (nonparasitic). The best known example is
the freshwater ciliate Paramecium (Figure 15.19e) .
Slime
Molds These protists are more attractive than their name. Slime molds
resemble fungi in appearance and lifestyle, but the similarities are due to
convergent evolution; slime molds and fungi are not at all closely related. The
filamentous body of a slime mold, like that of a fungus, is an adaptation that
increases exposure to the environment. This suits the role of these organisms
as decomposers. The two main groups of these protists are plasmodial slime
molds and cellular slime molds.
Plasmodial
slime molds are named for the feeding stage in their life cycle, an amoeboid
mass called a plasmodium (not to be confused with Plasmodium, the parasite that
causes malaria). You can find plasmodial slime molds among the leaf littler and
other decaying material on a forest floor, and you won't need a microscope to
see them. A plasmodium can measure several centimeters across, with its network
of fine filaments taking in bacteria and bits of dead organic matter amoeboid
style. Large as it is, the plasmodium is actually a single cell with many
nuclei (Figure 15.20).
Cellular
slime molds pose a semantic question about what it means to be an
individual organism. The feeding stage in the life cycle of a cellular slime
mold consists of solitary amoeboid cells. They function individually, using
their pseudopodia to feed on decaying organic matter. But when food is
depleted, the cells aggregate to form a slug-like colony that moves and
functions as a single unit (Figure 15.21 ).
Unicellular Algae Photosynthetic protists are called algae (singular, alga). Their chloroplasts support food chains in freshwater and marine ecosystems. Many unicellular algae are components of plankton (from the Greek planktos, wandering), the communities of organisms, mostly microscopic, that drift or swim weakly near the surfaces of ponds, lakes, and oceans. More specifically, planktonic algae are referred to as phytoplankton. We'll look at three groups of unicellular algae: dinoflagellates, diatoms, and green algae (a group that also includes colonial and truly multicellular species).

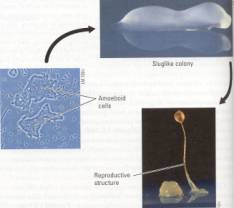
Left: Figure 15.20 A plasmodial slime mold. Pseudopodia of the
huge cell engulf small food particles in mulch or moist soil. The web-like form
is an adaptation that enlarges the organism's surface area, increasing its
contact with food, water, and oxygen. Within the fine channels of the
plasmodium, cytoplasm streams first one way and then the other, in pulses that
are beautiful to watch with a microscope. The cytoplasmic streaming helps
distribute nutrients and oxygen within the giant cell.
Right: Figure 15.21 Life cycle of a cellular slime mold. Most of
the time, cellular slime molds live as solitary amoeboid cells, using their
pseudopodia to creep through compost and engulf bacteria. When food is in short
supply, the amoeboid cells swarm together, forming a colony that looks and
moves like a slug. After wandering around for a short time, the colony extends
a stalk and develops into a multicellular reproductive structure.




Figure 15.22 Unicellular and colonial algae. (a) A
dinoflagellate, with its wall of protective plates. (b) A sample of diverse
diatoms, which have glassy walls. (c) Chlamydomonas, a unicellular green alga
with a pair of flagella. (d) Volvox, a colonial green alga.
Dinoflagellates
are abundant in the vast aquatic pastures of phytoplankton. Each dinoflagellate
species has a characteristic shape reinforced by external plates made of
cellulose (Figure 15.22a). The beating of two flagella in perpendicular grooves
produces the spinning movement for which these organisms are named (from the
Greek dinos, whirling). Dinoflagellate blooms-population explosions-sometimes
cause warm coastal waters to turn pinkish orange, a phenomenon known as a red
tide. Toxins produced by some red-tide dinoflagellates have caused massive fish
kills, especially in the tropics, and are poisonous to humans as well.
Diatoms
have glassy cell walls containing silica, the mineral used to make glass
(Figure 15.22b) .The cell wall consists of two halves that fit together like
the bottom and lid of a shoe box. Diatoms store their food , reserves in the
form of an oil that provides buoyancy, keeping diatoms floating as
phytoplankton near the sunlit surface. Massive accumulations of fossilized
diatoms make up thick sediments known as diatomaceous earth, which is mined for
its use as both a filtering material and an abrasive. Green algae are named for
their grass-green chloroplasts. Unicellular green algae flourish in most
freshwater lakes and ponds. Some species are flagellated (Figure 15.11c). The
green algal group also includes colonial forms, such as the Volvox in Figure
15.11d. Each Volvox colony is a ball of flagellated cells ( the small green
dots in the photo) that are very similar to certain unicellular green algae.
The balls within the balls in Figure 15.22d are "daughter" colonies
that will be released when the parent colonies rupture. Of all photosynthetic
protists, green algae are the most closely related to true plants.

Figure 15.23 The three major groups of seaweeds. (a) Green
algae. This sea lettuce is an edible species that inhabits the intertidal lone.
In addition to seaweeds, the green algal group includes unicellular and
colonial species, such as those in Figures 15.22c and d. (b) Red algae. These
seaweeds are most abundant in the warm coastal waters of the tropics. Of all
the seaweeds, red algae can generally live in the deepest water. Their
chloroplasts have special pigments that absorb the blue and green light that
penetrates best through
water. The species in this photo is an example of corraline algae,
which contribute to the architecture of some coral reefs. The cell walls are
hardened bya mineral. (c) Brown algae. This group includes the largest
seaweeds, known as kelp, which grow as marine "forests" in relatively
deep water beyond the intertidal lone. Some species grow to a length of over 60
m in a single season, the fastest linear growth of any organism. Kelp is a
renewable resource reaped by special boats that cut and collect the tops of the
algae. More importantly, kelp forests provide habitat for many animals,
including a great diversity of fishes. If you have walked on a beach covered
with kelp that has washed ashore after a storm, you may have noticed the organs
called floats, which keep the photosynthetic blades of the kelp in the light
near the water's surface. Maybe you even picked up and popped some of those
floats, the way you do those irresistible packing-material bubbles.
Seaweeds Defined as large, multicellular
marine algae, seaweeds grow on rocky shores and just offshore beyond the zone
of the pounding surf. Their cell walls have slimy and rubbery substances that
cushion their bodies against the agitation of the waves. Some seaweeds are as
large and complex as many plants. Even the word seaweed implies
plantlike appearance, but the similarities between these algae and true plants
are a consequence of convergent evolution. In fact, the closest relatives of
seaweeds are certain unicellular algae, which is why many biologists include
seaweeds with the protists. Seaweeds are classified into three different
groups, based partly on the types of pigments present in their chloroplasts:
green algae, red algae, and brown algae (Figure 15.23) .
Coastal people, particularly in Asia,
harvest seaweeds for food. For example, in Japan and Korea, some seaweed
species are ingredients in soups. Other seaweeds are used to wrap sushi. Marine
algae are rich in iodine and other essential minerals. However, much of their
organic material consists of unusual polysaccharides that humans cannot digest,
which prevents seaweeds from becoming staple food. They are ingested mostly for
their rich tastes and unusual textures. The gel-forming substances in the cell
walls of seaweeds are widely used as thickeners or such processed foods as
puddings, ice cream, and salad dressing. And the seaweed extract called agar
provides the gel forming base for the media microbiologists use to culture
bacteria in Petri dishes.
Evolution Connection
The
Origin of Multicellular Life
An orchestra can
playa greater variety of musical compositions than a violin soloist can. Put
simply, increased complexity makes more variations possible. Thus, the origin
of the eukaryotic cell led to an evolutionary radiation of new forms of life.
Unicellular protists, which are organized on the complex eukaryotic plan, are
much more diverse in form than the simpler prokaryotes. The evolution of
multicellular bodies broke through another threshold in structural
organization.

Figure 15.24 A model for the evolution of multicellular organisms
from unicellular protists.
(1) An ancestral colony may have formed, as colonial protests do
today, when a cell divided and its offspring remained attached to one another.
(2) The cells in the colony may have become somewhat specialized and
interdependent, with different cell types becoming more and more efficient at
performing specific, limited tasks. Cells that retained a flagellum may have
become specialized for locomotion, while others that lost their flagellum could
have assumed functions such as ingesting or synthesizing food. (3) Additional
specialization among the cells in the colony may have led to distinctions
between sex cells (gametes) and non-reproductive cells (somatic cells).
Multicellular organisms
are fundamentally different from unicellular ones. In a unicellular organism,
all of life's activities occur within a single cell. In contrast, a
multicellular organism has various specialized cells that perform different
functions and are dependent on each other. For example, some cells procure
food, while others transport materials or provide movement.
The evolutionary
links between unicellular and multicellular life were probably colonial forms,
in which unicellular protists stuck together as loose federations of
independent cells (Figure 15.2.4). The gradual transition from colonies to
truly multicellular organisms involved the cells becoming increasingly
interdependent as a division of labor evolved. We can see one level of
specialization and cooperation in the colonial green alga Volvox (see Figure
15.22d). Volvox produces gametes (sperm and ova), which depend on
nonreproductive cells, or somatic cells, while developing. Cells in truly
multicellular organisms are specialized for many more nonreproductive
functions, including feeding, waste disposal, gas exchange, and protection, to
name a few.
Multicellularity
evolved many times among the ancestral stock of protists, leading to new waves
of biological diversification. The diverse seaweeds are examples of the
descendants, and so are plants, fungi, and animals. In the next chapter, we II
trace the long evolutionary movement of plants and fungi onto land.